课题:2021-UNILausanne
主要内容:
通过施用抗冻蛋白与杀伤丁香假单胞菌来保护植物免受冻害
关键词:
抗冻蛋白;尾生菌素;CRISPR/Cas;丁香假单胞菌
1.问题发现与课题概述
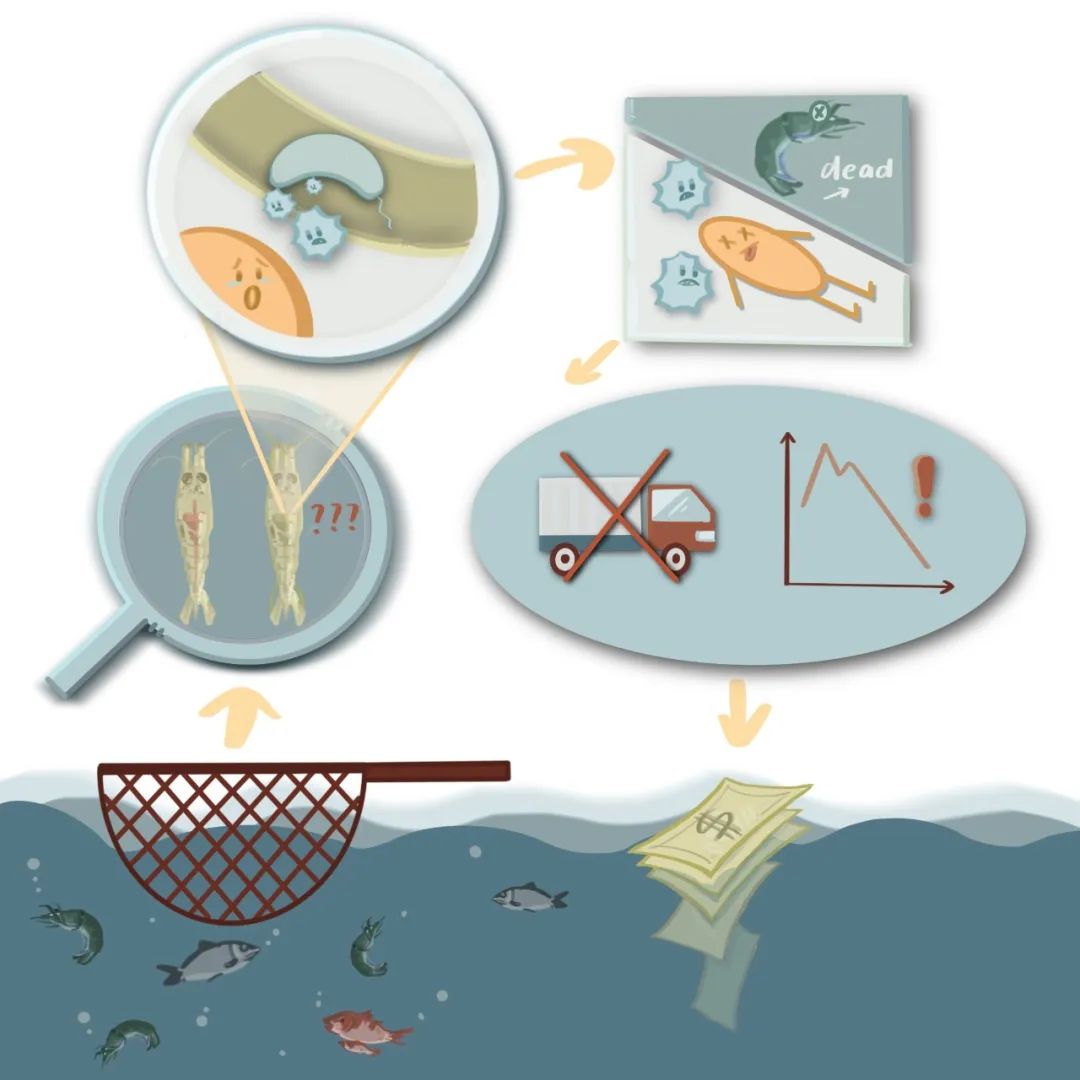
2021年的早春,该iGEM课题所在地经历了一次较为严重的霜冻:杏树的花苞被冻坏了,导致杏果的大面积减产。
本课题准备从两个方面来解决这一问题。其一,生产抗冻蛋白的保护液并施加给杏树,来保护杏树的敏感组织免受冰晶的损伤。其二,在冷冻期防治丁香假单胞菌的病原侵害。这种病原菌会形成冰核蛋白,促进冰晶的形成,增大霜冻期对杏树的危害;而从这一角度出发,本课题有表达植物尾生菌素和递送CRISPR/Cas9系统两种互补的方法。

图一
1.1抗冻蛋白
当温度降到零摄氏度以下的时候,冰晶破坏细胞,特别是花苞、幼年果实等敏感组织与器官。抗冻蛋白可以与冰晶结合,抑制冰晶的生长,从而减小冷冻期对植物的损害。
1.2噬菌体源性的尾生蛋白(Tailocin)
Tailocin是衍生自噬菌体的一种可以杀死竞争菌株的毒素,本课题中使用的Tailocin可以特异性地杀伤丁香假单胞菌,而不损伤其他的菌群。该类物质含有噬菌体的尾丝结构,它可以穿透植物的细胞膜,杀死靶细胞。由于这是噬菌体来源的,所以它具有比较高的特异性。同样的,我们仍然是利用大肠杆菌作为底盘菌来表达Tailocin。
1.3噬菌体
丁香假单胞菌的冰核蛋白是由InaZ基因编码的,如果将丁香假单胞菌的该基因沉默,那么就能从源头上阻止丁香假单胞菌产生冰核蛋白。本课题利用噬菌体来递送CRISPR/Cas9系统,并将该系统呈递给丁香假单胞菌。
1.4证明方法
课题组成员设计了一款能够准确调控温度的降温器,借助显微镜,可以监测样品的冷冻过程。除此之外,本课题组还设计了一套算法程序,来量化冷冻对植物的损伤。
2.抗冻蛋白详述与基因回路构建
抗冻蛋白的活性鉴定可以通过热滞现象(TH)或再结晶抑制活性测试(IRI)来验证。TH描述了非平衡冻结和融化温度之间的差异;IRI活性指的是防止冰晶继续长大的能力。不同的抗冻蛋白其所具有的活性是不一样的,细菌源性的抗冻蛋白具有适宜的TH和IRI活性,而来自鱼类或昆虫的抗冻蛋白有很强的TH活性,但是IRI活性却很低;来自植物的抗冻蛋白有很高的IRI,但其TH活性较低。本课题有三种候选的抗冻蛋白可供选择:松皮天牛、胡萝卜和黄杆菌的抗冻蛋白;抗冻蛋白在大肠杆菌中表达。

图二 抗冻蛋白的选择

图三 抗冻蛋白表达的质粒构建
关于质粒构建,本课题所采用的主要方法是用pET-17b和pCold-I两种不同的载体将其克隆到e.c oli BL21 (DE3)中,然后用IPTG诱导表达。最后利用磁珠、His-tag亲和柱和凝胶过滤对蛋白进行提取纯化。pET-17b含有一个T7启动子,可以调控重组蛋白的高水平表达。另一方面,pCold-I包括一个冷激蛋白a (cspA)启动子,该启动子允许较难表达和不能用T7系统表达的蛋白表达。因此,我们利用每个载体对AFPs的表达进行了比较,以优化其表达和合成。

图四 重组蛋白的少量表达与提取
3.噬菌体尾生菌素Tailoicins的选择与回路构建
植物的冻伤并不完全是物理因素造成的:有一些生活于植物表面的细菌种类, 它们能够促进冰晶的形成。在理想情况下,就算是易受寒害的植物都能够忍受-5℃至-10℃的冷冻侵袭,但是实际上,当温度稍稍低于0℃的时候,这些植物就会被冻伤,这就是由生存于它们表面的特殊的细菌导致的。在这些促进冰晶形成的细菌中,丁香假单胞菌就是一类典型的例子。
丁香假单胞菌可以产生一种衍生自噬菌体的细菌素:尾生菌素(Tailocins)。尾生菌素在结构上类似于噬菌体的尾管蛋白,是细菌用于菌际竞争的一类物质,具有较好的特异性,不会损伤其他的菌种。尾生菌素会结合在靶向细菌的细胞壁上,并在其上打孔,通过破坏氢离子梯度等方式来杀死靶细菌。

图五 尾生菌素的作用机制示意图
这种衍生自噬菌体的尾生菌素,相较于噬菌体本身具有一定的优势:
(1)宿主特异性(靶向性)较好,而且可以通过改变尾生菌素基因的靶向区域序列,来改变尾生菌素的靶向性;
(2)无法完成自我复制,如果被释放到环境中可以最大程度降低生物安全问题。尾生菌素的表达在诱导型启动子的基因回路中进行。
丁香假单胞菌有很多的菌株,杏树上的丁香假单胞菌是一类致病菌株。本课题组成员们无法确定杏树上生存的丁香假单胞菌是哪一类菌株,也没有找到对应的文献指明杏树上的丁香假单胞菌对哪一种尾生菌素敏感(这与我们副溶血性弧菌的课题的处境类似)。作为概念证明,本课题组选择了来自丁香假单胞菌DSM50252的尾生菌素,它可以靶向丁香假单胞菌B301D。这两种菌株都能够被买到(获得比较容易),二者的基因组测序信息也已知。B301D虽然最早是在梨树上被分离出来的,但是有文献表明它也曾经在杏树上被分离得到。
Tailocin基因是使用PHASTER鉴定的,这是一种能够鉴定细菌基因组中的噬菌体序列的工具。Tailocins含有原噬菌体的大部分元素,但缺乏整合酶和衣壳基因。所有的基因都存在于一个单一的Cluster中,其中包含了细胞裂解所需的结构基因和酶。结果显示,在我们的杀手菌株的基因组中同时存在一种原噬菌体和一种tailocin。Tailocin簇长13.8 kb,包含17个基因。有两种裂解酶:holin和溶菌酶。PHASTER将一些基因鉴定为结构尾基因;其中一种被鉴定为尾纤维,即负责目标识别和特异性的蛋白质。在尾纤维基因旁边还存在一种伴侣蛋白(尾纤维组装蛋白),帮助尾纤维正确折叠。最后,还有一些类似噬菌体的基因,其功能尚未被PHASTER(假设蛋白质)明确识别。

图六 尾生蛋白基因
接下来大致详述一下尾生菌素在底盘菌(大肠杆菌)中的生产过程。为了使尾生菌素的生产可控,本课题组成员在质粒构建中选择了诱导型的启动子(乳糖诱导型的T7启动子),所使用的质粒骨架是pSG3651Plibt7A。完整的尾生菌素基因簇是非常大的(13.8kb),所以本课题中将基因簇分成了四段,从菌体中PCR扩增下来,并采用Gibson组装的方法将片段连接在一起。

图七 回路组装示意图
4.噬菌体与CRISPR/Cas9方案
在本方案的最终目标里,课题组成员希望只沉默InaZ基因,而不去直接杀死丁香假单胞菌,最大程度上不破坏原生的微生物群。本课题中选用的是噬菌粒,这是一种类似于质粒的DNA,能够被包进缺陷型的噬菌体中。当噬菌体接触到丁香假单胞菌时,能够将噬菌粒注射进菌体,让噬菌粒发挥沉默基因的作用。

图八 噬菌体递送噬菌粒的示意图
在本课题组的系统中,Cas9蛋白产生之后就与sgRNA结合在一起(与靶标DNA分子结合)。当sgRNA靶向到了它的互补序列之后,Cas9蛋白会诱导一个双链的剪切,打开DNA双螺旋。

图九 CRISPR/Cas9基因敲除与同源重组修补
关于噬菌粒的构建与相关验证实验的设计,本课题将其划分为两个步骤。本课题并不直接验证基因工程改造的噬菌粒的效果,而是构建了与噬菌粒相统一的一套质粒系统,将质粒转化至B301D中,来验证CRISPR系统对InaZ基因的敲除效率。
【竞赛报名/项目咨询+微信:mollywei007】